|
 |
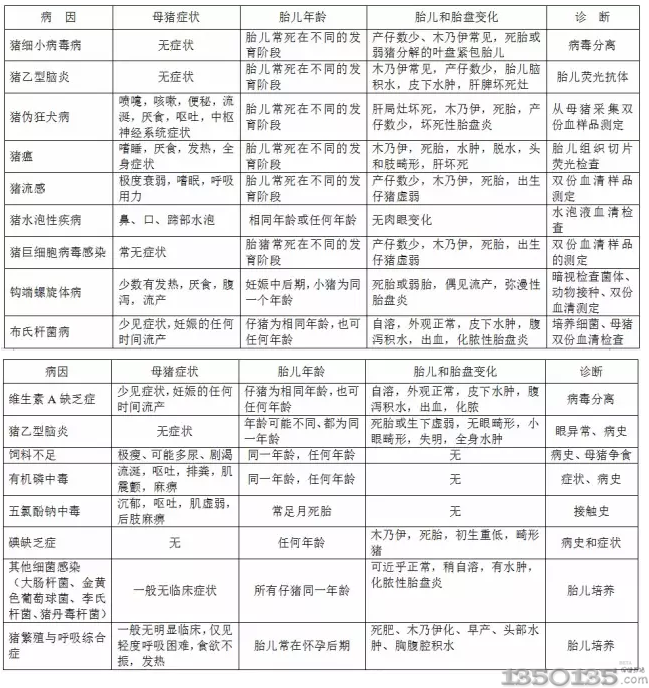
Ŀǰ�����i��Ѫ�U����Ⱦ�ѳɞ�ȫ���ԵĆ��}�����������ȣ��҇��яı��������������|�������ϡ����ϡ����ġ������ȵ^��һЩ�iȺ�з��x�����i��Ѫ�U����
�������i��Ѫ�U����Ⱦ�ɳʬF��N�R����ͣ����ͽ��^���߷Q��Glasser�ϲ���Glasser��s Disease�������w�S���Զ�l�ԝ{Ĥ�ס���l���P�����XĤ�מ����R���������еğo��l�ԝ{Ĥ��׃�����ʼ��Է����^��Ҳ�гʼ��Ԕ�Ѫ�Y���^�ġ��l�����g��2���g��4���g��ͨ����Ҋ��5��8���g�����i���l����r���õ��iȺ��һ���������������أ��������ױ��l��������Ⱦ�^������Ҳ������أ��l���ʺͲ����ʶ����ܸߣ���l���ʿ��_��10%��15%�������ʿ��_��50%�����i��Ѫ�U�����Ϻ������ij��������һ�N�C���²��������Բ��l���^�l���{�������ο�Ȯ������С���������A�h���������i�����߲���ʹ������s���������pʧ���ء����i��Ѫ�U����Ⱦ�c�i��Ĥ���ž��U�����i�����Ȳ�ԭ�wһ���ѽ���ͬ�ɞ鮔��F�������s�i���µ��韩���}֮һ��
����1 ���i��Ѫ�U��
����Glsser��1910���״Έ�������ڻ��Н{Һ���w�S������Ĥ�ס��İ��ס���Ĥ�ס��P�����XĤ�IJ��i�ĝB��Һ�аl�F���@�N���m����Լ����������x���B�ɹ����˿�����Schermer ��Ehrlich��1922����
�������о����ڣ�������Ҫ�����F߲�����͢��������������ʶ������ᣬNAD���ɷN���L���ӵ��i��Ѫ�U����Haemophilus suis�� ҕ��Glsser�ϲ����²����������Biberstein��White��1969���C������Glsser�ϲ��IJ�ԭ��ֻ��ҪNAD����ˣ��������h�Ѳ���Ҫ�a�oX���ӵ���Ѫ�U���������r��ǰ�Y“para������”��ʾ�^�e���Ķ����һ���·N——���i��Ѫ�U����Haemophilus parasuis����
����1.1�ΑB
�������i��Ѫ�U���ǰ��ϗU������Ѫ�U������һ�NС�U�������m����ԣ������\�ӣ����vĤ�ľ���һ�����U��o�vĤ�ľ����ΑB���ӣ��ʗU�λ�z�������ᡢ�Ӿ���x��ʮ���������廯����Cetavalone��������l�F���vĤ�к��ж�N�������|��Morozumi��Nicolet��1986�������u�߽që����Ĥ�����L�r������߀���γɽz���ΑB�����a����ë�ӽY��(Munch ��,1992)��
����1.2���B����������
�������������L��Ҫʹ�ú��Т����L���ӵ����B�����ɿ�����֬��Levinthal��֬����NAD��PPLO��֬�ȡ���ѪҺ��֬�ϱ����ڽ��Sɫ���Ѿ���̦��߅���l�Ǡ����L�����a�����ᣬ�l�������ǡ������ǡ���¶�ǡ����ǡ����Ǻ���ѿ�ǡ�
���������i���������x�����r�����ܕ����x����������Ѫ����ø��ԡ���هNAD����Ƽ������^ȥ�����@�������Ѫ�U��“СȺ��minorgroup��”һ�~���y�{������C��D��E��F֮�e������M������Ԕ�����������������Ɍ������c���i��Ѫ�U���^���_��D��E��FȺ���Ϻ������г��ڵľ�Ⱥ����Ҳ���ԏķλ��X�M���з��x����Moller���ˣ�1993����DNAͬԴ�Է����Ļ��A��ָ����D��EȺ����ͬһ�N���ʌ��@�ɂ�Ⱥ�ϲ����S���о������룬Moller�ȣ�1996������������·N��������“minor group”��D+EȺ��F Ⱥ���@3���·N�քe��С�ž��U����Actinobacillus minor�����i�ž��U����A.porcinus��������ž��U����A.indolicus���������Ё��Ժ�������V������ه�ļ����M��16SrRNA���б��^������l�F�����i��Ѫ�U���c����ž��U����FȺ���Pϵ������У�ͬԴ���_��97.4%��97.7%�����ߵIJ�e�ǣ�����ž��U�����Ԯa������Ͱl�������ǡ�
����1.3Ѫ����
����Ѫ������о��Y�����������i��Ѫ�U��������Ŀ�ԭ�Ծ��и߶Ȯ�Դ�ԡ�Bakos�ȣ�1952����������ԇ�ĽY��������A��B��C��D�Ă�Ѫ���ͣ�֮��Morozumi��Nicolet�����7��Ѫ���ͣ�1��7����Kiestein��(1991)��������6����Jena6��Jena12����Kielstein��Rapp��Gabrielson��1992�����b����5����Ѫ���ͣ�ND1��ND5��������ʹѪ����͵õ��yһ�� Kiestein��Rapp��Gabrielson��1992���������خ����ÿ�Ѫ���M�е����ߔUɢԇ�Y����������i����Ѫ�U���µ�Ѫ���ͷ���wϵ���������^ȥ���C����Ѫ����1��7��Ѫ����Jena��ND�ϲ���Ѫ����8��15��Ŀǰ���@һ���ͷ����ѱ������ɼ{�������_���i��Ѫ�U���F��15��Ѫ���ͣ�1��15����ֵ��һ����ǣ��M����ˣ�߀���д����ķ��x�겻�ܱ����͡�
�S�������о��˱���Ѫ���͵ķֲ���r�����ձ��������������������������ô���Ї�����Ѫ��4�͞郞���ͣ�Ѫ��5��Ҳ�������x��(Morikoshi�ȣ�1990��Kielstein��Rapp-Gabrielson��1992��Rubies�ȣ�1999����Tadjine�ȣ�2004��Cai�ȣ�2005)���Ĵ������͵������е���Ѫ��5��13�ͣ�Blackall�ȣ�1996, 1997��Rafiee��Blackall��2000��Angen��2004����
����1.4 �����Ͷ�������
�������i��Ѫ�U���Ķ�������������δ���塣Ѫ��W����ͨ��Ҫ���]���c�������Pϵ����Ѫ��5��10��12��13��14�;��꽛��ǻ;����ȾSPF�i����l���Ͳ����߷弯����4d֮�ȣ�����J�飬��������Ѫ���ͣ�Ѫ��2��4��15�������l�ԝ{Ĥ�ף��o���������ж����ͣ�ʣ�µ�Ѫ���ͣ�3��6��7��8��9��11���������κ��R���Y���o��Ѫ���ͣ�Kielstein and Rapp-Gabrielson, 1992��Amano ��,1994�����IJ��i��������������λ���x�����꣬Ѫ���̈́e���ƣ���Ѫ��2��4��5��12��13��14���͡������^Ѫ�������Р�B���{��Y�����������������ⲿλ���x���ľ��꣬Ѫ������Ҫ��1��2��4��5��12��13��14���Լ��S��߀���ܷ��͵ġ����Խ����i�Ϻ������ģ���Ҫ��Ѫ��3�ͺͲ��ܷ��͵ġ�
����������֪�����ϗU���ƳɆT����һЩ�dz���Ҫ�Ķ����������vĤ����ë��֬���ǣ�LPS������Ĥ����(OMP)�ȡ����ǣ����i��Ѫ�U���Ƿ�ͮa���@Щ�������ӣ��Լ������c�����Ķ����Ƿ����P��Ŀǰ߀֪֮���٣��˴�ì��֮̎�H�ࡣ
���������\���˹���Ⱦԇ��о����vĤ�c�������Pϵ����Little��Harding��1971���Լ�Morozumi��Nicolet��1986��������Ľ����i��ǻ�Ͳ��i�����Ы@�õľ��꣬���vĤ�IJ����ࡣ
����Munch�ȣ�1992���C�������i��Ѫ�U�����w�Ȃ������܉����ɾ�ë�ӽY���������²����ò������
����LPS������һ����Ҫ�Ķ������ӡ����ǣ�Zucker�ȣ�1996����δ�l�F�������LPS�c�o�����LPS�����@�IJ��ͬ�ӣ�Miniats�ȣ�1991���l�F���ú���LPS��OMP��ԭ�ľ������߄���H����OMP���w���i���ֿܵ��������Ĺ������@Щ���ƺ�������LPS���Ǹ���Ѫ�U������Ҫ�������ӡ�֮��Amano�ȣ�1997���Mһ���о���LPS���²����ã�����Ѫ��5�;���ӷN�����l�F��Ѫ����LPS���w�ij��F�cѪ˨�γɺ͏�����Ѫ�܃���Ѫ���P���F����������i��Ѫ�U����LPS�����c�������m����Լ����ȶ������ƵĻ��ԣ�Raetz��Whitfield��2002����
��������SDS-PAGE���g�l�F�����i��Ѫ�U������Ĥ����(OMP)����2����ȫ��ͬ�������ͣ�����1�ͺ�����2�͡��ɽ����i��ճĤ���x���ľ�����_����1��OMP���������һ��s68Kda����һ��23��40KDa֮�g����Glsser�ϲ����i���x���ľ���ͨ��������2��OMP�����Լs37KDa�ă��ݵ��מ������������Oliveira��Pijoan��2004��ͨ�^ȫ�����M��Ӌ��C�����C�����@Щ�l�F��
�������i��Ѫ�U������һ���������ӿ���������ø��Lichtensteiger��Vimr��1997���l�F��90������Ұ����a������ø��ԓø�ڱ����Č������L��ĩ���_ʼ���_������ø��������ͨ�^ø�����¶�鱾����ֲ������������������������w�������ķ���ϵ�yҲ����ճ������Զ���Ɖġ�
�����c�������P�Ķ��أ�������δ�l�F��Schaller�ȣ�2000��Ҳ�ų��˸��i��Ѫ�U�����Юa���c��Ĥ���ž��U��RTX��Apx���������P�Ķ��صĻ���
����Blackall�ȣ�1997���\�ö���λø�Ӿ��multilocus enzyme electrophoresis��MEE�����gԇ�D����������������x���c���������x��֮�g�ą^�e���Y���ǣ��������H��ʾ�˲�ͬ��Դ�����g�ľ�ͬ�c�����Ұl�F��ͬһѪ���Ͳ�ͬ�����g�ľ��e���о��ˆT�������֞�2��MEE��Ⱥ�����ǣ�Ҳ�]�аl�F������x��λ�cMEEȺ֮�g���Pϵ��
����Hill�ȣ�2003���\�òRT-PCR���g(differential display RT-PCR)�о��˾���1185��Ѫ��5�ͣ��Ķ�����ԓ�����c���Բ������Ƶ�40��ߜؗl�������L�����߰l�F��7�������ڱ��_�������քe�cfadD��֬�������oøA�ϳ�ø����apaH�������������ᣩ��pstI�������D��ø�wϵø��cysK�����װ���ϳ�ø����StD��Na����Cl-��ه�x���D�\���ף���HSPG������������Ĥ�خ��ĸ��������о����ǰ�w����PntB����ऺ����D��ø������ͬԴ��15��Ѫ���Ͷ�����ͬ�ı��_�������c�������ӵ��Pϵ�����о��С�
��1.5 ��������W����
����������͵ķ����ѱ����ڸ��i��Ѫ�U������ķ��������cѪ����ͷ����^���������^���@��ͨ�^�b������DNA�D�V��l�F��ijЩDNA�D�V�����c�������P������ȫ����Ⱦ���x���ľ����c���²��Ծ����Ȼ��ͬ��ǰ�߸������gͬԴ���ஔ�ߣ�Oliveira��2003����
����Smart�ȣ�1988�����������ԃ���øָ�y�D�V����REE���о���SPF�iȺ�ͳ�Ҏ�iȺ���i��Ѫ�U���ķֲ���r���ڽ^��SPF�iȺ�������l�F���ƈD�V�ľ��꣬����Ҏ�iȺ����ĈD�Vʮ�֏��s���l�������i�w���x���ľ��꣬��D�V�����ƣ����cͬһ�iȺ�ɽ����i��ǻ���x���ľ���D�V��ͬ��
���������؏�Ԫ����PCR��repetitive element based��PCR��rep-PCR�����gҲ�����ڸ��i��Ѫ�U���Ļ�����͡�ԓ����ͨ�^ERIC��enterobacterial repetitive intergenic consensus������U����С��ͬ��DNAƬ�Σ��ٽ��Ӿ���x����ʾ�خ�����M�D�V�����@�N�����M�з�������ʹ����ͬѪ���͵ľ���Ҳ�ɷֱ����ͬ��DNA�D�V���о��l�F�����������i�ľ��겢���࣬���������������Ƶ�DNA�D�V��Versalovic��,1991,1994��Woods��1993��Rafiee��,2000��Oliveira��,2003����
�����M�и��i��Ѫ�U��������͵���һ�N������PCR������ø���Ӿ�L�ȶ��B��ԇPCR��restriction fragment length polymorphism ��RELP�����\���@�N��������tbpA��һ�N���a�\�F���Y�ϵ��Ļ�����15��Ѫ���͵Ę˜ʾ���ֳ���12����ͬ��DNA�D�V������Ѫ��5��12��14��15�͘˜ʾ���������Զ��B�D�V��ͬ�����b��101��Ұ����r���l�F��33��RELP�D�V��������10���c�˜ʾ������ͬ��δ�l�FѪ���ͺ�RELP�V���g�����P�ԣ�Redondo�ȣ�2003����
����2 �l���C��
�������i��Ѫ�U������Ķ�ֲ��λ�Ƿ�����Ϻ������������Пo��Փ��Vahle�ȣ�1995�����Ǹ�Ⱦ5���gCDCD���i���l�F�ӷN��36h�ɏ�ѪҺ����ǻ�͚�܃ȷ��x���ӷN��ڷκ�ѪҺͿƬ�в��װl�F������Ҳ�]�Џı����w�з��x����Amano�ȣ�1994����Ѫ��1��4��5�;��꽛�ǽӷN���i�t�ı�ǻ�ͱ����w�з��x�����i��Ѫ�U����Segales�ȣ�1997������ښ�܃ȽӷN���ɏı����w�͚�������з��x���ӷN�2001��Kirkwood�Ȅt����ˏ�Ⱦ�i��ǻ�����з��x�����i��Ѫ�U���ĽY����
������ɸ��i��Ѫ�U���ּ�ȫ�������ӣ���δ����ҹ��J��Vahle�ȣ�1997���C�����ı�ǻ�жη��x�����i��Ѫ�U�����i�����м��Ի�ē�Ա����Һ�wë������mucocilliary cell����ʧ������߀������@ЩճĤ�ĸ�׃�����ڸ��i��Ѫ�U���������Լ����_Ѫ�������ǣ��oՓ��ͨ�^��R߀��ͨ�^���߽M����������δ�����wë��ʧ��ճĤ����׃�ԵIJ�λ�ҵ����i��Ѫ�U����
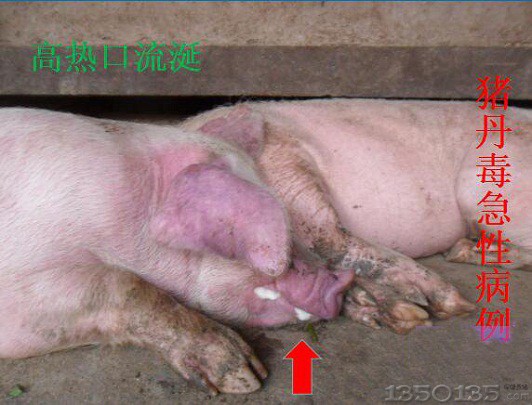
����Brockmeier��2004���C����֧��ܔ�Ѫ���ϗU����B.bronchiseptica������ɸ��i��Ѫ�U�����Ϻ�������ֲ���T���������c�iή�s�Ա����ж������ϗU�����������ơ�
����3 ����W
�������i��Ѫ�U����Ⱦ�ʵط������С���Ҫͨ�^ֱ�ӽ��|������������ĸ�Ⱦ���iȺ���M�����i�����M�²��Ծ��ꡣ�����g�i����Ⱦ�����ɱ��l������ͬ�ӣ�һ����ԭ�Բ�ͬ���µď������������iȺ��Ҳ���������߲����l��Oliveira��Pijoan��2002����
�����ڸ�Ⱦ���iȺ�У�ĸ�i�Dz����ă������oՓ���²���߀�Ƿ��²��Ծ��꣬���i�����ڲ����ڱ���Ⱦ�ġ��������ϣ�ĸ�i�Ď����ʲ����ߣ���˱���Ⱦ�����iҲֻ��һС���֡�����Ⱦ��С�i�����Ⱦ���a�������������Ժ�Ҳ���ܳɞ遆�R�������i��δ���²��Ծ����Ⱦ�����i���ɏ�ĸԴ���w�@�ñ��o����5~6���g���̕r��ĸԴ���wˮƽ�½�������������ص�Ӱ푶���ʹ���R�������i�������ӣ���Щ�ڲ����ڛ]�н��|�^�²��Ծ�����iֻ�_ʼ�l������ˣ��������R���Y��ͨ����5~6���g�r������֮����F��
����4 �R���Y��
�������i��Ѫ�U����Ⱦ����F���R���Y���c��Ⱦ��λ���P��Hoefling��1994������֞�4�N��ͣ���Glsser�ϲ����w�S���Զ�l�ԝ{Ĥ�ף�����Ѫ�Y�ͣ��o��l�ԝ{Ĥ�ף������Լ����ͣ�myositis acuta ��Ҋ��ҧ�����ͺ������͡�
�������R�����F����^��������Ƿ��خ��Եġ����̳���Ի��Խ��^���l���iͨ����ʮ�ֽ��ѵ��i������İY�����w�����ߡ����������ʳ���p�ˣ��������F���ԡ��������y���w���½������С�����ʧ�{���l�C���P�ز���ȰY��еIJ��i˥�߶�����
��������l���ļ����Ͳ����кܶ�N������Ĥ���ž��U�����i�ž��U�����i�������i�����U�����i��֧ԭ�w�����в����ȡ������@Щ��Ҫͨ�^�b�e�\������ų���
����5 �\��
�������i��Ѫ�U����Ⱦ���R���Y����خ��ԣ�����R���\�����x���_�\������Ҫ�M�Ќ���ҙz
����5��1 ����׃��
������������Ҫ׃�����ڸ�Ĥ���İ�����Ĥ���P������F�{Һ-�w�S���Ի��w�S���Ի�ē���װY���M���W�z���@Щ�װY��λ��Ҋ������Ѫ�����;��ȼ��������������������XĤ�ס�˨�����XĤ�X�ף������X��Һ���ࡣ�β���׃�����͡���Ѫ�Y�������ڸΡ��I���X�����ٱ���Ҋ����Ѫ�ߣ��c����ͬ�rѪ�{�Ѓȶ��غ������ӣ������ٳ��F�w�S�����K�����аl�C��Ƥ��ˮ�[�ͷ�ˮ�[�ļ��Ԕ�Ѫ�Բ�������Ҋ��ҲҊ�������͵ĝ{Ĥ�װY���F��
����5��2 ��ԭ���x
�����IJ��Ϸ��x���i��Ѫ�U����ͨ���ڽӷN�˽��Sɫ���������ѪҺ��֬���M�У�Ŀ����ͨ�^���Sɫ��������鸱�i��Ѫ�U�������L�ṩNAD��Ҳ�����x���ɿ�����֬������NAD��PPLO���B�������B�r�gͨ����Ҫ24~48h���������y�����B������֮һ�������ǏIJ����з��x�r�������ܵ��s���ĸɔ_����Q���k����һ�Dz���ϡ����B�������������B���м��ֿ�ù�غͿ����ġ����ڱ������Ϻ��������ھ�����ˣ����ע���ɺ��������x����������һ�������l����ȫ���Ը�Ⱦ�����ǣ������X���P���șz�����i��Ѫ�U�������\���rֵ�Ͳ��������ˡ����⣬�����x�����i��Ѫ�U���Ӽ����r����회��������µ������b������֮�c��������Ѫ��NAD��ه����������ž��U�����i�ž��U����С�ž��U���ȅ^�e�_��
����5��3 Ѫ�嶨��
�����������������i��Ѫ�U����Ѫ�����ѽ������خ����ÿ�Ѫ�����ߔUɢԇ�ĽY�����_����15��Ѫ���ͣ�1~15���������߀���ˌ���ͬ��Ѫ��W���ͷ���[���ߔUɢԇ�(ID)���g��Ѫ��ԇIHA���ͅfͬ����ԇCA��]�M���˱��^�о���Río�ȣ�2003��Tadjine�ȣ�2004��Turni��Blackall��2005����
����Turni��Blakall(2005)���^��ID��IHA�z�yҰ����ĽY����l�F��IHA�a����ID����IJ��ɷ��;��꣬ǰ���_44�������ߞ�41����Del Rio�ȣ�2003����Tadjine�ȣ�2004�����о��t�J�飬���m�ϸ��i��Ѫ�U��Ѫ����͵ķ�����ԓ��IHA�����IHA�p��Ѫ��W���ɷ��͵İٷ֔����ٱ�ID��CA��10���������W��Ԕ��������Del Rio��Tadjine�ȵ��о��Y����l�F���������о�������ȱ�ݣ������z�y�ľ���]��һ���LJ��H�J�ɵĘ˜ʾ��꣬�����õĿ�ԭ��ȡ���gҲ�]�н�GDԇ�У������ˣ��������о��Y���ܵ��|�ɡ�
�������x���겻�ɶ��͵�ԭ���������������خ���ԭ�Ĕ������㣬Ҳ������������δ���J֪��Ѫ�����͡�ID��IHA���ͽY�����F���ԭ�����������õĿ�������Ȼ��ԭ��������ID�ġ���飬IHAԇ����ÿ����Կ�ԭ�����tѪ��������Y���ǰѳ�����ԭ��������ԭʹ�ã��Ķ�ʹIHAԇ�������Ա�IDԇ������3000����Mittla��2003����
����5.4 ���w�z�y
�����a�w�Y��ԇCF�����g��Ѫ����IHA��ԇ��ELISAԇ�����ڙz�y���i��Ѫ�U���Ŀ��w�����Բ������^һ�ܵIJ��̼��əz�����w���������^���@���ͽ��淴���ԡ�Takahashi�ȣ�2001���������߽ӷNԇ��ͨ�^CFԇ�z�y���w�ζȣ���Y���ڶ����19d�ʬF��Եζȡ��о��l�F���oՓ���ó��۴��߀���üӟ���еľ��w�������d��t�����Ŀ�ԭ�M��IHA���oՓ������м���������Һ߀���÷ӣ�ˮ���������Һ����ԭ�M��ELISA����ֻ�ܫ@�ò���������ԽY�����e���ڙz�y���߄���r������f���@Щԇ�����ڙz�y���߱��o���������ǣ�������ø����R�֜���ȫ���w����ԭ�M��ELISAԇ�t�������о�ĸ�i�Ŀ��w�ζȺ����i������ӷN������
5.5 ��������W����
�����@����У�һ�����Б���ǰ���ķ����ǹѺ����خ��Բ�ƽ���s��ԇOSCPH��oligonucleotide-specific capture plate hybridization�����������Ժܸߣ���102CFU/mL�����B����}�Dz����@�ü����B���ˣ��ֽ�����һ�N��ֱ���b�������и��i��Ѫ�U����PCR�����������������_102CFU/mL�����ң��������ڙz�y�ќ��IJ�����Ȼ�����@�ɷN����������ž��U�������ԳʬF����Է������ɴˣ�Oliveira��Pijoan��2004�����]�@�ɷN����ֻ���ڙz�y����������ĸ��i��Ѫ�U�����������ž��U�����Ϻ������еij��ھ���
����5.6���߽M���\��
���������s�����L��ʹ���i��Ѫ�U�����x�����D�y������������]�����߽M������IHC�����\�ั�i��Ѫ�U����Ⱦ��IHC�ă��c�ǣ���ʹ���i��Ѫ�U���ѱ����ɼ���������z���ڰ��{�У�Ҳ���p�ر��z�y����
�������}�DZ���������ʹ�õĶ��¡���w�c��Ĥ���ž��U���н��淴����Segales�ȣ�1997����
����6 �����A��
�����c��������һ�ӣ��l�����ӡ��I�B���m�����������T���Դ��M���i��Ѫ�U����Ⱦ�İl�������С��e�ǟo���DȺ���Լ���ͬһȦ����B��ͬ���g���iֻ�����Ǵٳɱ������l���T��
����6.1 ��������
�������i��Ѫ�U���������ÿ������ί����ί�Ҫ��M��M�죬�Y��һ�����F������θ�c��;���oˎ����ˎ������Y����������Glsser�ϲ������ڲ�ԭ�M��M�����X��Һ�Լ��ֺ��P������ˎ����һ��Ҫ��
������ʲô�����أ���ԓ����ˎ��ԇ�Y�����x�����P������ˎ�ԵĈ�����ࡢ��ʿ���x�������и��i��Ѫ�U�����ꌦ��ù�غͶ���ɳ�Ǿ����У����ù�ء�����ù�ء��c��ù�ء��ĭh�ء��tù�ء��ǰ���ऺ�TMP+�ǰ���श��п��ԣ�wissing�ȣ�2001�����������ꌦ���y����ˎ��(���S��ù�ء��^���渣���h��ɳ�ǡ��tù�ء������ῼ����ù�ء����^ù�ء��ĭh�ء���ճ���ء������ǡ�TMP+�ǰ���������)��ʮ�����У�Aarestrup�ȣ�2004����
����6.2 �����A��������ӷN
����ͨ�^����ӷN�����Ƹ��i��Ѫ�U����Ⱦ�����ܱ��^����Щ���mȻ�������Ķ������Ӻͱ��o�Կ�ԭ����߀����������_���������خ��Ե���������������������ɏS��ُ�I��Ҳ���������ԼҾ��硣���ԼҾ�����Ƽ������^���У���Ҫ���]��������Ѫ���ͣ�ҲҪ��ҕ���������������ӡ�����һ�^�i���Է��x���������o���ĸ��N���ꡣ�ɴ����]�����ʹ�����X�ȷ��x���ľ�����Ƃ��ԼҾ��磻��t������Ï��P�������w������λ���x���ľ������磬����Ч����׃����÷Ⱦ����Ƃ�t�����oЧ����ԭ����ǰ��������ͬ��λ��Դ�ľ����зdz��ߵĮ�Դ�ԣ�heterogencity����
�������ڸ��i��Ѫ�U��Ѫ���͵Ķ����Ժʹ�����IJ��ܶ��͵ķ��x��Ĵ��ڣ���ʹ�˂���Ŭ�������܉�a�����汣�o�������硣Miniats�ȣ�1991������ԇ�Dʹ�ú��и��²�������²�������ľ�����T�����汣�o�����߷������Y���ǣ��ֿ�ͬԴ�Ժͮ�Դ�Ծ���Ľ��汣�o����ֻ���ö��������Ƃ�ľ�����ܵõ������²��������Ƃ�ľ���ӷN���i���H�ܱ��oͬԴ����Ĺ�����Rapp-Gabrielson�ȣ�1997�����о�Ѫ��2��4��5��12��13��14���g�Ľ��汣�o�ԕr�l�F������Ѫ��12���Ƃ�Ćξ���Ѫ��2��12���Ƃ�Ķ��r���⣬�����Ͷ��܌�ͬԴ����a�����o����Ѫ��4���Ƃ�ľ��磬���Ա��oѪ��5�͵Ĺ�������Ѫ��4��5���Ƃ�Ķ��r���磬���Եֿ�Ѫ��13��14�͵Ĺ��������i��׃�ć����ԺͲ��������@���ͣ���ԓ����߀�Mһ���о���Ѫ��12�;��猦ͬԴ����ı��o�Է�����Ѫ��12����12a��12b�ɂ����ͣ��քe��֮�Ƃ����������i�l�F����12a�Ƃ�ľ��粻�ܮa��ͬԴ�Ա��o������12b���ߣ��t�ܮa���@����ͬԴ�Ա��o���@Щ���������M��Ѫ��12�;��궼�Ǹ��²��Եģ�Ҳ�ܮa�����Ƶ�OMP��LPS���������������_�ı��o�Կ�ԭ�϶���������ͬ�ġ�Takahashi�ȣ�2001���о���Ѫ��2��5��֮�g�Ľ��汣�o�ԣ��C���r�������߲��ܮa�����汣�o����������r�������ߣ�Ȼ������������2��5�;���քe�������p�p���ܫ@�ñ��o��Bak��Riising��2002���о���Ѫ��5��+DiluvacForte����ı��o�ԣ����i��5���g��7 ���g�ӷN��������a���ı��o�����H�ֿܵ�Ѫ��5��ͬԴ����Ĺ��������Ҍ�Ѫ��1��12��13��14 �͵Ȯ�Դ����Ĺ���Ҳ���ܮa�����@�ı��o���á�
�����A�����i�l�����������iС�����ظ�Ⱦ�²��Ծ��ꡣԓ��������Փ���A�ǻ����@��һ�N���O������Ȼ��B�£��iȺ�к������i��ֲ���²��Ծ��ꣻ�������ڶ�ֲ�����i�����i���H������ĸԴ���w����lȫ���Ը�Ⱦ����ĸԴ���w�p�ٕr���i�ܫ@���������߷���������ͨ�^���gԇ��C�����@�N���O�������i��5���g�r���ڽӷN7×103CFU�����������Ⱦ�i�IJ����ʱȌ����i������2.88%����Ҫ��ӛ����Ⱦ��PRRS�������i����ʹ���@�N������
��������ӷN����һ�����}������r�C���}���ЌW�߲�ͬ������ӷNӰ��Ԅ����ߵ��^�c�������C������ĸ�i�����i�ӷN����ͬ����Ч���J����鿹�w���ܸɔ_����Ч����Ҳ���˳ֲ�ͬ�^�c�����ĸ�iʲô�����ӷN��ֻ�����i�ӷN���磬�Y���oЧ����ˣ����˱��C���i����ǰ�͔��̺��܌�����ӷN�a�����߷������б�Ҫ��ҕ�OӋ����ӷN�IJ��ԡ�Oliveira��Pijoan(2002)�C����ĸ�i����ǰ���߮a������֭�����Լ��������i������ӷN���ٽӷN���܉��ṩ��Ҫ�ı��o���á�
����7 �YՓ�c���h
�������^ȥ�Ĕ����У��mȻ�˂�������ҕ�ˌ����i��Ѫ�U����Ⱦ���о��������������S�����}���ܵõ��M��Ļش��e�nj��������ӵęz�y���������ӵ����ÙC�����\����ĸ��M�����͏V�V������аl�ȣ���Ҫ������ҕ�����о���
�������҇�����עҕPRRS������PR�������i�A�h�������i��С�����������ͬ�r����ԓע����i��Ѫ�U���IJ��l���^�l��Ⱦ���̓������á������ڼ��s���̶ȺܸߵĴ����i����������ȡ�����Ľ���Ӗ��������ҕ���i��Ѫ�U���ĝ���Σ���ԡ��������о��^���У�Ҫ�����҇��i�������������ԣ�Ҫ�������Ƹ��i��Ѫ�U�����r�磬߀Ҫ�_ʼ�����A����N�������硣